The field of mammalian genome engineering took a huge step forward with the birth of dolly the sheep and the advent of cloning technology 22 years ago. At the time it was hoped that cloning would provide us with the ability to introduce precise insertions or deletions easily and efficiently into the germline of livestock species. Unfortunately, as it turns out, cloning is not as easy nor as efficient as we had originally hoped. The rodent community have developed stem cells which have allowed them to bring about improvements in the efficiency of cloning. The same cells do not exist for livestock, and as such, the livestock community has had to look elsewhere for improvements in genome modification.
Ironically, the first tool in the genome editors’ tool-box, the zinc finger nuclease [1], was first described in the same year that Dolly was born. Since then a number of other editor reagents have been developed, namely the TALEN in 2010 [2] and more recently the CRISPR/Cas system in 2012 [3]. These tools allow us to introduce precise insertions or deletions at far higher efficiencies that has even been possible before.
Genome editors are essentially DNA scissors that generate a double strand break (DSB), they can be utilised in a number of ways, however, their predominant use is to generate a targeted DSB. How that break is subsequently repaired allows the user to introduce changes at the target site (see Figure 1).
Figure 1
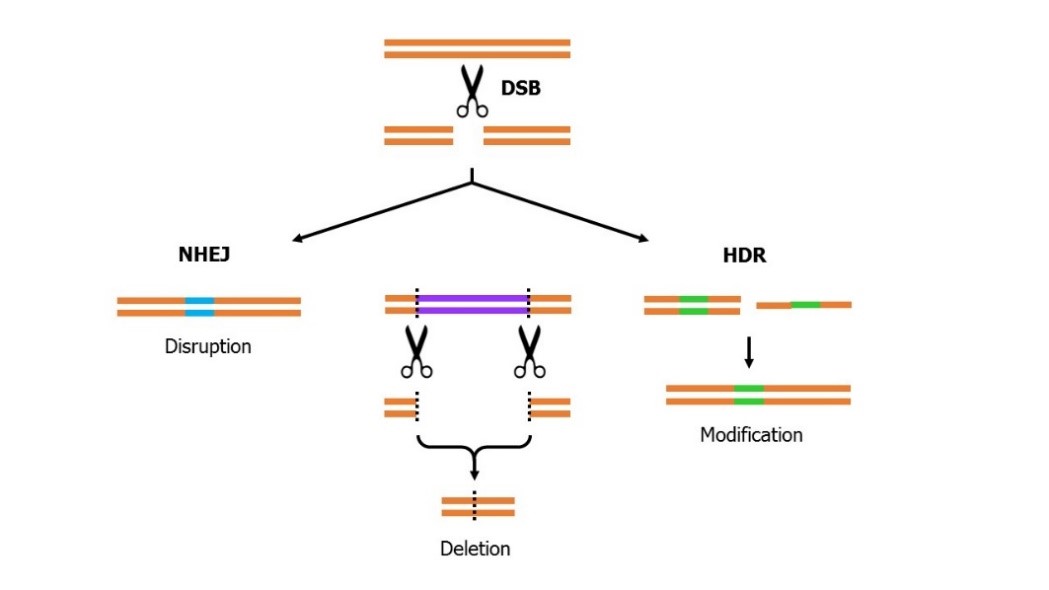
If the cell is allowed to repair the break itself it predominantly utilises the non-homologous end joining (NHEJ) repair pathway which is an error prone process often leading to the introduction of small insertions and/or deletions (indels) at the site of repair. These indels can result in a frameshift leading to premature termination of the protein and loss of function. NHEJ, in that respect, is an ideal approach for analysis of gene function as it allows researchers to knock out a gene. Alternatively, to introduce specific changes at the site of the DSB it is possible to bias the cell towards the alternative homology dependant repair (HDR) pathway. To achieve this a template for repair of the break is supplied, either as a single strand oligonucleotide for small changes or as a double stranded DNA molecule, typically incorporated into a plasmid, for larger changes. The template acts as a scaffold for repair of the break resulting in introduction of the changes encoded by the template at the site of repair. Finally, genome editors can be utilised to remove an undesirable region of the genome you could use a pair of genome editors. To do this genome editors can be designed to generate DBSs flanking the region to be removed, with repair of the resulting breaks leading to deletion of the intervening sequence. It is important to note that genome editors do not allow us to do anything that we couldn’t do before, however, it significantly increases the efficiency of doing so meaning that the conversation changes from ‘if’ to ‘when’.
In terms of genome editing in livestock, most of the work carried out still utilises the cloning technology forged with Dolly but in this case using somatic cells that have been edited to generate animals containing the desired genome changes.
Certainly, if you were wanting to make large genome changes or introduce transgenes this would be the way to go. At The Roslin Institute, however, we prefer a more direct approach, where we simply inject the editing reagents in to the zygotes of livestock species before transferring them to recipients in order to generate the desired edited animals and we have had great results using this technique.
One question you may have at this point is ‘why use genome editing technology in breeding systems?’ The simple answer is that it allows you the ability to import traits, more specifically, traits that are currently difficult to introgress through conventional breeding whilst maintaining genetic merit. Specific examples of this are the introduction of traits from out-with the breeding population, traits from a different species or the removal of deleterious traits that are fixed in your population. Indeed, there are a number of published examples of genome editing being successfully applied in cattle to improve production, welfare and disease resistance, highlighting the potential of this technique.
The first example of genome editing in cattle was a modification of the myostatin gene, which is a negative regulator of muscle growth. Belgian Blue cattle have an 11 bp deletion in the myostatin gene which leads to a non-functional protein and an increase in muscle mass known as the ‘double muscle’ phenotype. The aim of this project was as a proof of principle for the technology showing that you could introduce the double muscle phenotype in to cattle that do not naturally possess the trait. In addition, the advantage of choosing the myostatin gene as a target is that it requires the creation of the simplest of the genome edits to produce the desired effect, as the generation of an indel at the target site is likely to result in a loss of functional protein and an increase in muscle mass. For this project genome editing reagents were designed to generate a DSB in the genome of Nelore cattle in approximately the same region of the gene as the natural deletion in the Belgian Blue cattle. Two live born animals were generated, a heterozygous edited bull and a wild type heifer. The edited bull had a similar deletion to the one observed in the Belgian blue and resulted in a clear increase in muscle mass when compared to its wild type sibling [4]. This result not only validated the technology but also prompted researchers to target other traits requiring more specific manipulation of the genome.
One such trait was polled (hornless) cattle. In the United States, it is estimated that 80% of all dairy calves (4.8 million per year) and 25% (8.75 million animals) of all beef cattle are dehorned every year. The dehorning of cattle is not only a costly process but it is also painful for the animals. The practice has recently come under the spotlight due to public concerns about animal welfare with a number of groups campaigning for the enforcement of anesthesia during dehorning or the banning of dehorning as a farming practice. Introduction of the polled allele into the Holstein population has been attempted by cross breeding with polled breeds, such as Angus. Unfortunately there is a significant cost to the estimated breeding value of the progeny of such crosses due to the significant reduction in milk production, the effects of which would take numerous generations to rectify. Genome editing, however, has been used to generate polled dairy cattle in a single generation with no impact on milk production [5].
Improving disease resistance is a significant challenge facing the farming industry today especially due to the requirement to produce more food for an increasing global population. Genome editing tools are ideally placed to aid this and have been used to tackle a number of critical diseases.
Bovine tuberculosis (TB) is a devastating chronic disease of cattle caused by Mycobacterium bovis (M. bovis) and has a major impact on the cattle industry. Genetic studies have shown that cattle with natural resistance-associated macrophage protein-1 (NRAMP1) variants show differing resistance to M. bovis. Genome editors have been used to change the NRAMP1 gene to variants associated with higher resistance to TB and cells isolated from the generated cattle show increased resistance [6].
The three examples I have discussed so far have used genome editors to either knock out protein function or to modify existing proteins. As yet there has been no example of an undesirable region being deleted from the genome in cattle but has in pigs when addressing the issue of porcine reproductive and respiratory syndrome (PRRS). PRRS is a global disease of pigs that causes abortion in sows, death of new born piglets and a failure to thrive in older animals. It is the most economically important disease of the pig industry, and annual production losses are estimated to cost the industry €1.5bn in the EU. It is also important to note that vaccination strategies attempted have thus far proved ineffective. The PRRS virus infects macrophages, and gains access to the cell via interaction with the macrophage-specific surface protein CD163. Furthermore, we know that this interaction is limited to domain 5 of the protein, and that domain 5 is encoded exclusively by exon 7 of the gene. We designed genome editors that flanked exon 7 to delete it from the genome. Using alveolar macrophages isolated from the resultant animals we showed that the remaining protein was still folded correctly, trafficked to the cell surface and retained the activity associated with relevant cellular functions. Critically CD163 no longer acted as a viable binding partner for the virus making the cells totally resistant to infection [7].
Genome editors have a huge potential to make a significant impact on the livestock industry, however, there are a number of issues that still need to be clarified before the technology is likely to be more widely applied. We need to remember that the technology is still in its infancy and we need to not get carried away. Yes genome editors have fantastic potential, but as scientists we are still learning about the technology and the possibilities and we need to work with breeding companies to determine how it would operate in a breeding system. Licensing issues need to be resolved, most notably with respect to the CRISPR/Cas9 system where there are ongoing patent battles. Perhaps the biggest question is how will this technology be regulated? There are two sides to this issue; the lack of movement on the regulation of genome editing and the quagmire that is the current regulation of genetically modified organisms.
In terms of the regulation of genome editors, the UK Government says its policy will be in line with the evidence in this area and it broadly supports the view that genome modification (GM) and genome editing (GE) are safe. In a report on advanced genetic techniques for crop improvement, the House of Commons Science and Technology Committee itself stated ‘we are convinced by the evidence provided to us that this suite of technologies is a potentially important tool, particularly in the developing world, which should not be rejected unless there is solid scientific evidence those technologies may cause harm’.
The European Academies’ Science Advisory Council published a report stating that ‘livestock breeding should also be governed by the same principle as proposed for plant breeding – to regulate the trait rather than the technology’ (EASAC policy report 31) but the report didn’t confirm whether organisms produced by genome editing will be subject to the existing GM controls but there has been no date given for this. The interpretation of the EU definition of what constitutes a regulated GM organism, however, is not straightforward. As a result, the councils report will no doubt prompt further discussions as to the appropriate approach to be followed at EU level. In the US the FDA regulate intentional DNA changes in animals as drugs (FDA draft revised guidance #187) whereas the USDA have decided that some genome edited plants fall out with its regulatory remit [8]. As such there is a push for both organisations to employ a more harmonised approach.
So, where does that leave us with genome editors? One of the big questions is how will genome editors be classified – as a novel breeding technology or an advanced GM technology? Potentially it might come down to what the edit is, whether it be the introduction of a SNP or a larger change. However, genome editors can also be used to improve the efficiency of transgenesis. So should it be the animal that is regulated and not the technology? Ultimately genome editors, in a lot of cases, simply accelerate the breeding process.
Whatever happens we need to make sure that the first GE animal to go through the system is the right model for the right reasons. Meaning that there are clear benefits in terms of animal welfare, the environment and to the consumer. The first animal through the system will likely be used as a test bed and set the bar for future animals. We need to make sure that the science behind the model is well considered. Due diligence would dictate that we check for unintended consequences of the editing. Taking the PRRS example we need to make sure that by making pigs that are resistant to PRRS we haven’t inadvertently made them more susceptible to other diseases.
Finally, we have an extremely powerful tool that has opened up a huge wealth of possibilities and has the potential to have a massive impact on the livestock industry. If we are to be able to utilise it to the fullest we need to have a system where the regulations are based on the science and not politics.
There are three types of editing event that can be created utilising genome editor generated DSBs. Non-homologous end joining which often leads to indels at the site of repair which can result in gene disruption. The homology dependant repair pathway allows the introduction of modifications at the site of repair by providing a single strand or a double stranded DNA molecule that acts as a scaffold for repair of the break. Genome editors can also remove an undesirable region of the genome by generating DBSs flanking the region, the repair of which leads to deletion of the intervening sequence.
References
1. Kim, Y.G., Cha, J. and Chandrasegaran, S. (1996). Hybrid restriction enzymes: zinc finger fusions to Fok I cleavage domain. Proc Natl Acad Sci USA, 93(3): 1156–60.
2. Christian, M., et al. (2010). Targeting DNA double-strand breaks with TAL effector nucleases. Genetics, 186(2): 757–61.
3. Jinek, M., et al. (2012). A programmable dual- RNA-guided DNA endonuclease in adaptive bacterial immunity. Science, 337(6096): 816–21.
4. Proudfoot, C., et al. (2015). Genome edited sheep and cattle. Transgenic Res, 24(1): 147–53.
5. Carlson, D.F., et al. (2016). Production of hornless dairy cattle from genome-edited cell lines. Nat Biotechnol, 34(5): 479–81.
6. Gao, Y., et al. (2017). Single Cas9 nickase induced generation of NRAMP1 knockin cattle with reduced off-target effects. Genome Biol, 18(1): 13.
7. Burkard, C., et al. (2017). Precision engineering for PRRSV resistance in pigs: Macrophages from genome edited pigs lacking CD163 SRCR5 domain are fully resistant to both PRRSV genotypes while maintaining biological function. PLoS Pathog, 13(2): e1006206.
8. Ledford, H. (2016). Gene-editing surges as US rethinks regulations. Nature, 532(7598): 158–9.